





Huseníček jako modelová rostlina
Od roku 1990 se v mnoha zemích intenzivně pracuje na výzkumu genomu huseníčku. Projekt se oficiálně jmenuje Multinational Coordinated Arabidopsis thaliana Genome Research Project. Zahrnuje jednak sekvencování celého genomu, jednak studium funkce všech genů a jejich hlavních interakcí. Jeho cílem je porozumět na molekulární úrovni principům uspořádání a funkce rostlinného genomu, růstu, vývoji, fyziologii a biochemii této modelové rostliny a po zobecnění i ostatních vyšších rostlin.
Sekvencování genomu A. thaliana by mělo být dokončeno r. 2004. Roku 1996 bylo založeno konsorcium AGI (Arabidopsis Genome Initiative), sdružující laboratoře z Evropy, USA a Japonska, které jsou společnými silami schopny sekvencovat desítky milionů párů bází ročně. K tomu, aby byl metodou náhodných úseků zachycen celý genom, bude potřeba asi sedminásobná redundance v sekvencování. Zatím bylo sekvencováno přibližně 60 milionů párů bází DNA, tedy přibližně polovina genomu.
 Druhým důležitým aspektem je studium funkce všech genů. Běžně se vychází z mutací získaných chemickou a radiační cestou, ale v poslední době je nejvýznamnější transpozonová mutageneze a mutageneze prostřednictvím plazmidu T-DNA Ti-bakterie Agrobacterium tumefaciens. Tento přírodní systém vnášení genů z bakterií do rostlin je základem techniky vytváření většiny transgenních rostlin a lze jej využít také k vytvoření mutací nového typu. T-DNA vyvolává svým začleněním do genů rostlinného genomu mutace a zároveň slouží jako značkovací úseky ke klonování mutovaných genů. K tomu, aby bylo s 95% pravděpodobností postiženo všech 20 000 předpokládaných genů huseníčku, je třeba vybudit asi 100 000 nezávislých inzercí. Brzo bude i tento úkol splněn co se týče počtu inzercí, ale ne co se týče počtu mapovaných a sekvencovaných genů. Všechny získané linie s inzercemi T-DNA, i ty, u nichž mutace běžnými metodami zatím zjištěny nebyly, se uchovávají jako cenný zdroj informací pro další fázi studia.
Druhým důležitým aspektem je studium funkce všech genů. Běžně se vychází z mutací získaných chemickou a radiační cestou, ale v poslední době je nejvýznamnější transpozonová mutageneze a mutageneze prostřednictvím plazmidu T-DNA Ti-bakterie Agrobacterium tumefaciens. Tento přírodní systém vnášení genů z bakterií do rostlin je základem techniky vytváření většiny transgenních rostlin a lze jej využít také k vytvoření mutací nového typu. T-DNA vyvolává svým začleněním do genů rostlinného genomu mutace a zároveň slouží jako značkovací úseky ke klonování mutovaných genů. K tomu, aby bylo s 95% pravděpodobností postiženo všech 20 000 předpokládaných genů huseníčku, je třeba vybudit asi 100 000 nezávislých inzercí. Brzo bude i tento úkol splněn co se týče počtu inzercí, ale ne co se týče počtu mapovaných a sekvencovaných genů. Všechny získané linie s inzercemi T-DNA, i ty, u nichž mutace běžnými metodami zatím zjištěny nebyly, se uchovávají jako cenný zdroj informací pro další fázi studia.
Ke klonování genů lze využít i mutace získané chemickou a radiační mutagenezí, postup je však zdlouhavější. Předpokládá velmi podrobné chromozomální mapy. Využívají se při tom knihovny umělých chromozomů kvasinek (YAC) nebo bakterií (BAC).
Již byly dokončeny fyzikální mapy (založené na délce DNA jednotlivých chromozomů a jejich fragmentů po štěpení restrikčními endonukleázami) všech pěti chromozomů. Potvrzuje se, že hustota genů je asi 1 gen na 5 kilobází DNA. Zjištěno a analyzováno bylo přes 400 genů. Asi 90 % je zcela nových a výskyt řady z nich se u rostlin nepředpokládal. U organizmu s tak malým genomem je značně překvapivý výskyt mnoha genů ve formě multigenových rodin. Obvykle však geny nejsou uspořádány ve funkčně příbuzných skupinách, často se skládají z neobvykle malých exonů (překládaných úseků) a malých intronů (nepřekládaných úseků, které jsou eliminovány na úrovni RNA). Vazbová mapa (mapa pořadí genů a molekulárních znaků) huseníčku v současné době zahrnuje více než 500 lokalizovaných genů a téměř 1000 sond DNA. Kromě toho je k dispozici asi 30 000 krátkých sekvencí bází komplementárních k nalezeným mRNA (EST). Takto je podchycena mRNA asi 10 000 genů, z nichž některé lze určit pomocí několika EST. To jsou většinou geny, jejichž transkripce nastane jen výjimečně – ve specializovaných buňkách nebo pouze za zvláštních podmínek. Značná část z nich jsou pravděpodobně regulační geny a předpokládá se, že je bude možno detegovat inzerční mutagenezí.
Součástí klonování genu je důkaz totožnosti klonovaného genu tím, že se zpětně vnese do genomu huseníčku, v němž byl původní gen zbaven aktivity. Takto vnesený transgen obnoví ztracenou funkci.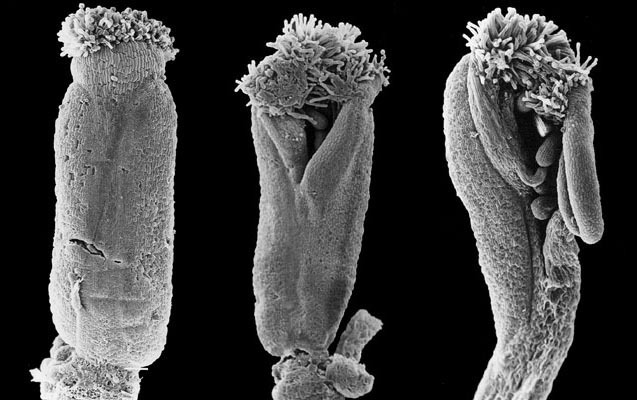
Důležitou součástí studia genomu huseníčku je srovnávací mapování. Porovnává se struktura celého genomu i jednotlivých genů s genomy a geny příbuzných i vzdálených rostlinných druhů. Je tu velká podobnost například s druhy rodu Brassica. Ukazuje se, že pořadí genů zůstává v krátkých blocích zachováno i u obilovin.
Předpokládá se, že po dokončení hlavní etapy studia genomu huseníčku budou geny této modelové rostliny (buď v původní formě, nebo různým způsobem upravené) používány ke zlepšení genomu kulturních rostlin. Krom toho tato modelová rostlina poslouží jako vzorová a její geny budou využívány jako sondy k vyhledávání homologních genů v jiných rostlinných druzích. Z teoretického hlediska bude možno studovat vztahy v signalizačních drahách regulace genetické informace na novém základě, vyplývajícím z jednoznačné znalosti pořadí bází DNA v genech a pořadí aminokyselin v proteinech.
Studium genomu A. thaliana přineslo zatím většinou teoretické poznatky zásadního významu. Praktické uplatnění transgenů A. thaliana, i když je stále v počátcích, již rovněž přineslo první nadějné výsledky. Stále častěji kupříkladu slouží jako základní zdroj genů pro zlepšování genomu (a potažmo tedy i vlastností) kulturních rostlin. Vybrané příklady jsou uvedeny v rámečcích vložených do textu.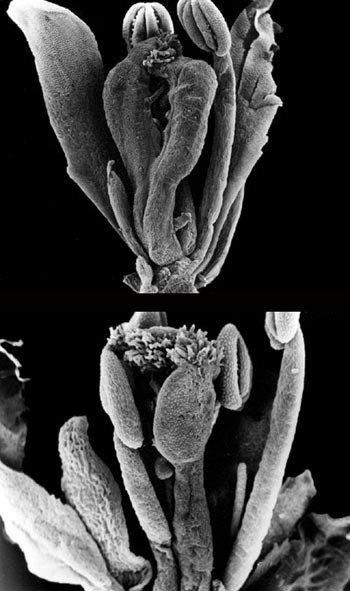
Transgen pro odolnost nízkým teplotám
V loňském roce byla v Kanadě uvolněna k polním pokusům transgenní vinná réva se zvýšenou odolností proti mrazu. Do jejího genomu je včleněn gen A. thaliana pro enzym superoxiddismutázu. Superoxiddismutázy katalyzují přeměnu superoxidových radikálů (02–) aktivovaného kyslíku na peroxid vodíku a kyslík. Tyto radikály vznikají v rostlinných buňkách ve zvýšené míře při stresových podmínkách, jako je mráz, sucho či zaplavení. Rostliny mají superoxiddismutázy, ale přidání dalších genů vede někdy ke zvýšení odolnosti k chladu. Zvýšení tolerance k chladu bylo dosaženo také u vojtěšky po transgenozi tímto genem a předpokládá se, že se transgen uplatní v mnoha dalších druzích kulturních rostlin.
GEN PRO INICIACI TVORBY KVĚTŮ
Jeden z genů iniciace kvetení huseníčku, označovaný leafy (LFY), byl klonován a přenesen do genomu osiky. Tam podmiňoval kvetení semenáčků již prvním rokem, přestože osika za normálních okolností kvete až za 8–20 let. Tento gen je přítomen u všech dvouděložných rostlin. Vnesení dalšího genu do rostlinného genomu bude pravděpodobně snižovat minimální dobu vegetativního stadia, potřebnou k dosažení stadia generativního. Mechanizmus genetické regulace kvetení byl u huseníčku popsán již dost podrobně. Jde o síť vzájemného působení genů a je velký úspěch, že se podařilo najít rozhodující gen.
Vrcholový meristém (vzrostný vrchol) A. thaliana má tři diferenciační programy, které umožňují různé utváření základních opakujících se článků rostlinného těla – segmentů nodů s listy a internodií lodyhy:
- Prvním typem diferenciačního programu je tvorba přízemní listové růžice. Každý segment má velmi krátké internodium, list růžice a úžlabní pupen, který obsahuje meristém, z něhož se může vyvinout další lodyha. Listová růžice tvoří bazální část lodyhy květenství.
- Druhý typ tvoří bazální internodia květenství. Segment má dlouhá internodia, list růžice a úžlabní meristém. Z úžlabního meristému se může vyvinout lodyha s dalším stupněm úžlabních pupenů. Květenství Arabidopsis thaliana je složené racemické květenství, charakteristické pro celou čeleď Brassicaceae.
- Třetí typ segmentů jsou segmenty vlastního květenství v horní části lodyhy. Každý segment má internodium střední délky zakončené květem, a list s úžlabním pupenem mu chybí.
Existence genu Leafy byla poznána na základě homeotické květní mutace, v níž všechny větve primárního a sekundárního květenství tvoří vedlejší květenství. Pokud vzniknou terminální květy, pak se skládají jen z kališních lístků a plodolistů. Gen byl klonován a zjistilo se, že kóduje transkripční faktor, který působí pozitivní regulaci – mění meristém květenství na květní meristém. Přenesení původního nemutovaného genu transgenozí do genomů jiných rostlin podmiňuje ranější vznik květů.
Produkce hirudinu
Geny A. thaliana se již více způsoby uplatnily při transgenozi řepky olejné. Do genomu řepky olejné byl například včleněn chimérický gen, jehož kódující sekvence byla složena z kódujících sekvencí genů pro dva různé proteiny. První část byla z genu pro speciální protein semen A. thaliana a řepky – oleosin – a druhá část byla z kódující sekvence genu pro hirudin pijavky koňské (Haemopis sanguisuga). Pijavky při sání krve vylučují do rány ve slinách protein hirudin a ten zabraňuje srážení krve. Tento protein by se uplatnil při komplikovaných operacích k prevenci embolií, pokud by ho byl dostatek. Protein může být syntetizován v semenech transgenní řepky olejné, kde fúzní protein oleosin-hirudin tvoří až 1% transportních proteinů. Hodnota jednoroční sklizně na poli, osetém takovouto transgenní řepkou, pak několikanásobně převyšuje hodnotu pole. Oleosiny (i fúzní protein) jsou vysoce lipofilní proteiny, které se hromadí na povrchových membránách olejových tělísek v buňkách děloh semen řepky. Lze je snadno oddělit od zbytku buněčných extraktů centrifugací.
Existuje ještě řada dalších typů farmakologicky významných vzácných proteinů, o kterých se předpokládá, že je bude možno produkovat transgenními rostlinami. Není pochyb o tom, že upravené geny A. thaliana se uplatní při transgenozi rostlin mnoha způsoby buď přímo, nebo jako sondy pro nalezení homologních genů jiných organizmů, jež pak budou po dalších úpravách využívány k transgenozi.
Plevel A. thaliana, který se stal modelovou rostlinou, tak slouží jako základní zdroj genů pro zlepšování genomu kulturních rostlin transgenozí.
Ke stažení
 Článek ve formátu PDF [351,69 kB]
Článek ve formátu PDF [351,69 kB]
O autorech
Doporučujeme

Divocí kopytníci pečují o krajinu

Relativistický čas – čas našeho světa

